Licence: Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0)
Credit: Fungal genetics / J.R.S. Fincham and P.R. Day. Source: Wellcome Collection.
25/334
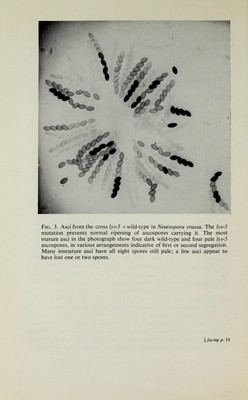
![THE CHROMOSOME THEORY 11 anything between zero and about 67 per cent and, as will be explained below, can be used as a measure of the distance of the locus of the allelic difference from the centromere of the chromosome. The characteristic behaviour of segregating alleles can be shown most graphically where the genetic difference affects the appearance of the ascospore itself. Several such cases are known in different species of Ascomycetes ; an ex¬ ample from Neurospora is illustrated in Fig. 3. Independent assortment and linkage When the diploid nucleus of the ascus initial contains a pair of unhke alleles, such as the A-a pair determining mating type, it is said to be heterozygous with respect to the chromosomal locus in question. When two identical alleles are present the ascus is said to be homozygous. The mating type locus is the only one for which Neurospora asci are invari¬ ably heterozygous, but a great number of allelic differences at other loci are available for experiment, mainly as a result of artificially induced mutation (see Chapter 3). It is thus possible to study the types and arrangements of ascospores obtained from asci heterozygous at two or more loci simultaneously. As an example we will take a sample of 1161 asci dissected from a single cross by Howe [188]. The parents of this cross actually carried different alleles at four different loci, but for the moment we will restrict our attention to two of them. One parent was of mating type A and required riboflavin for growth because of the presence of the allele r, while the a parent had only the wild-type nutritional re¬ quirements and carried the corresponding wild-type allele r+. It may be noted that the + superscript is generally used to denote alleles norm¬ ally present in the wild-type. The data for the mating type and r loci are shown in Table 2. The main point which emerges from Table 2 is that the segregation of A-a is independent of that of r-r+. It will be seen that the frequency of second division segregation of r-r+ is even lower than that shown by mating type (2 per cent as compared with 12-5 per cent). Consequently the most frequent classes of asci are those in which both allelic pairs segregated at the first division. Among such asci it seems to be a matter of chance whether r is segregated with A or with a at the first division. The difference between the frequencies of the two cases (473 as against 521) is hardly significant statistically (the appropriate statistical test for significance, the test, is outUned in Chapter 4). The frequencies of the other classes of asci are also reasonably close to what would be expected, given the frequencies of second division segregation of the two loci, on](https://iiif.wellcomecollection.org/image/B18023691_0026.JP2/full/800%2C/0/default.jpg)





 No text description is available for this image
No text description is available for this image No text description is available for this image
No text description is available for this image No text description is available for this image
No text description is available for this image