Genética y metabolismo / Robert P. Wagner [y] Herschel K. Mitchell ; traducción del inglés por Faustino Cordón.
- Wagner, Robert P.
- Date:
- 1963
Licence: Attribution-NonCommercial 4.0 International (CC BY-NC 4.0)
Credit: Genética y metabolismo / Robert P. Wagner [y] Herschel K. Mitchell ; traducción del inglés por Faustino Cordón. Source: Wellcome Collection.
29/556 (page 25)
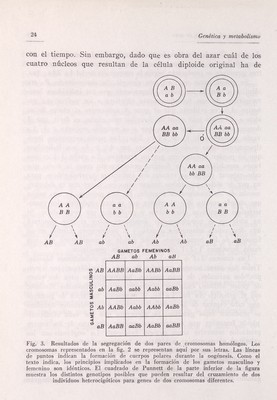
![Introducción al concepto de gene 25 ser el que reciba el citoplasma, los mismos tipos de gametos cabe esperar en la oogénesis que en la espermatogénesis, siempre que los genotipos de las células germinales diploides sean los mismos. El factor esencial que hay que tener presente en toda meiosis que implique más de un par de homólogos es que es cuestión de puro azar que los no homólogos se acompañen uno a otro en la primera división de la meiosis. Así, en la fig. 3, en una célula con cromo¬ somas marcados АаВЪ, A puede acompañar a В y, por consiguien¬ te, a irse con Ъ ; o A y b pueden ir a un polo, y, por tanto, a y В al polo opuesto. Por consiguiente, como se ha señalado antes, cabe esperar cuatro tipos de gametos de una célula heterocigótica para dos pares de genes de cromosomas diferentes. Tres pares de genes alélicos sobre otros tantos cromosomas diferentes darán origen a ocho gametos diferentes. De la fecundación entre gametos de los genotipos señalados en la figura 3 resultan los genotipos que señala el cuadrado de Pun- nett. La razón genotipica es, sin duda, compleja por el gran nú¬ mero de genotipos que se obtienen. El tipo de razón fenotípica dependerá de los tipos de relaciones de dominancia entre los aleles y también de posibles interacciones entre los genes no alélicos. No consideramos ahora tales interacciones, pero se coinsiderarán por extenso en el capítulo X. Si se supone una dominancia com¬ pleta de Л y В sobre sus respectivos aleles y que no hay interac¬ ción de genes, resultará una razón fenotípica 9:3:3:1. Esto pue¬ de escribirse QAB ] ЗАЬ :3aB -Aab, puesto que AB puede conside¬ rarse que es la designación fenotípica de АаВЪ, АаВВ, AABb, А ABB, у del mismo modo para los demás. Otros tipos de razones que resultan de las interacciones de genes se dan en la tabla 36 del capítulo X. El hecho de que estas razones puedan deducirse de la razón más sencilla 3 :1 obtenida cuando sólo se segrega un par de genes alélicos, puede apreciarse por la aplicación de alguna probabilidad simple. Por ejemplo, en el cruzamiento de A a x Aa, las probabi¬ lidades de que un descendiente posea el gene A (es decir, de que sea bien Aa, o bien AA) son de 3 entre 4, ó 3/4. Hay la misma probabilidad de que posea В un descendiente de un cruzamiento](https://iiif.wellcomecollection.org/image/b18033933_0030.JP2/full/800%2C/0/default.jpg)